Birusak bizidunen eta bizigabeen arteko mugan kokatu izan ohi dira. Izan ere, birioiek ez dute berezko makinaria metabolikorik, hau da, ez dira aurrekari metabolikoetatik hasita beraien osagai estrukturalak beren kasa eraikitzeko gai. Horregatik, ugaldu ahal izateko, beste izaki batzuk (ostalariak) inbaditu eta beroien zeluletako makinaria metabolikoez jabetu behar dute.
Birusek bere barnean babesturik daramaten informazio genetikoaren baitan, zelula infektatuaren metabolismoa inhibitu eta birusaren osagaien sintesia bideratzeko aginduak daude metaturik. Beraz, birusak parasito molekularrak dira, hainbatetan, harrapari bihurtu eta kutsaturiko izakia gupidarik gabe heriotzera eraman dezaketenak.
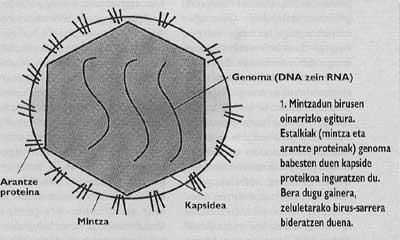
Beraz, erreplikatzeko , birusek ezinbestekoa dute beraien genoma (ADN zein ARN) ostalariaren barnera nola edo hala sartzea. Horretarako jarraitu behar duten prozesuari sarrera deritzo. lnfekzioa gertatu ahal izateko sarrera derrigor burutu behar denez, horrek garrantzi handia du birusen bizi-zikloan. Zelula eukariotikoetan sartzen diren birusen kasuan, birioiek ezinbestekoa dute zelula inguratzen duen kanpo-estalki molekularra edo mintza gainditzea.
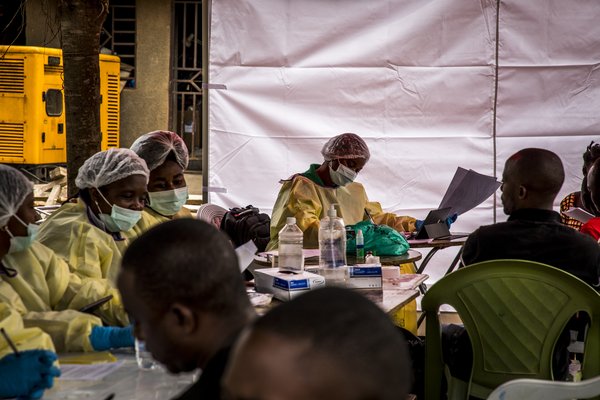
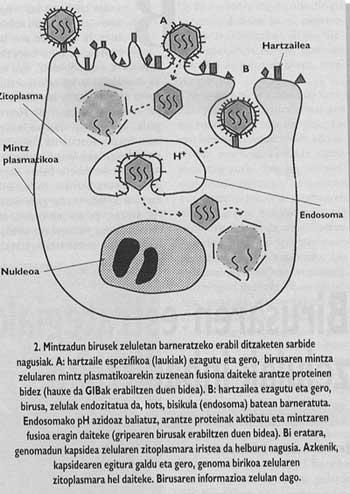
Halaber, animaliak eta gizakia infektatzeko gaitasuna duten birusik inportanteenak estalki deritzon zelulena bezalako mintz batez inguratuta egoten dira (1. irudia). Sarrera bideratzeko beraz, birus horiek beren estalki propioak inbadituko dituzten zelulen estalkiekin bat egiteko mekanismoak garatu dituzte. Helburu-zelularen eta birioiaren mintzak bat egitea lortuz gero, birusaren genoma zelularen zitoplasmara hel daiteke (2. irudia) eta bertatik erreplikazioa bideratzen hasi.
Birus estalkidunek garatu dituzten sarrera-estrategiak oso prozesu konplexuak dira eta horiek ondo ulertzeko, zientzilariak etengabeko ahaleginak egiten ari dira, etsai erahiltzailearen sarrera nola ekidin ikasterik bagenu, hainbat infekziorako babesa lortzeko bidean egongo ginateke eta. Infekzio birikoak inhibitzeko modu sinplea eta orokorra aurkitzeko asmoz, ikerlariak sarreraren elementu amankomunak bilatzen saiatu dira.
Tamalez, gauzak ez dira geuk nahi bezain sinpleak. Orain arte dakigunez: behintzat, birus-familia bakoitzak sarrera-estrategia desberdina garatu bide du eta ondorioz, sarrera-mekanismo orokor eta amankomuna dagoenaren hipotesia indarra galduz joan da. Gainera, birusen sarrera-mekanismoei buruzko ikerketek oraindik ez dute emaitza askorik eman, 80ko hamarkadaren hasieran (zientzilariak gripea sortzen duen birusaren aurkako txerto unibertsala ekoizterik ez zegoela jabetu zirenean, hain zuzen) garatzen hasi baitzen alor hori eta oraindik sortu berritzat jo daiteke.
Espainiako INIAko Rafael Blasco ikerlariaren hitzetan: "edozein birusi buruzko ikerketa hilik dago horrek sortutako gaixotasuna tratatzeko txertoa garatzen den unean". Blasco jauna nafarreria sortzen duen Poxvirus -aren moduko birusen sarreran estatuan dagoen espezialistarik garrantzitsuena dugu. Nafarreriaren birusa, gizakiok planetatik nahita desagertarazi dugun lehen izakia omen da eta ondorioz, garrantzi gutxikotzat hartzendiren bere familiakideen sarrerari buruz ardura gehiegirik ez dagoela dio Blascok.
Planteamendu horren akatsa agerian geratu da azken bolada honetan, gripearen eta HIESaren izurriteak nola gelditu ez dakigularik. Gaur egun ezinezkoa zaigu hurrengo izurritea eragingo duen birusa zein motakoa izango den aurrikustea. Ezintasun hori gainditzeko bidean, aurrerapauso garrantzitsua izango litzateke edozein infekzio biriko tratatzeko baliogarri izango litzatekeen terapia unibertsala garatzea. Horretarako halaber, nahitaezkoa da birusen sarrera-prozesuak sakonki ikertzea. Gripearen birusa, adibidez, geure babes inmunitarioaz trufatzen espezialista da. Birusak eta antzeko beste agente arrotzak ere neutralizatu ahal izateko, agente arrotz horiek ezagutu eta desberdintzen aldez aurretik "ikasi" behar dute giza gorputzaren defentsek. Hori, txertoen bidez lortzen da gaur egun. Txertoek agerian jartzen diete gorputzeko defentsei agente arrotzen identitatea eta horrela, agente horiekin bat egin behar duten antigorputzak ekoiztu beharko dituzten zelulak infekzioa gertatu baino lehen prest egotea lortzen da.
Gripearen kasuan ordea, gauzak ez dira hain errazak. Gripearen aurkako txertoak urtero aldatu behar izaten dira, gripearen birusaren zepa bakoitzak antigeno desberdinak (hau da, antigorputzek identifikatu ahal dituzten molekula birikoen talde desberdinak) izaten baititu. Are gehiago, birusak berak hurrengo belaunaldietan aldatu egingo zaizkiola jakinik molekula horiek agerian nahita jartzen dituela dirudi. Horrela, erraz sorraraz ditzakegu birusaren aurkako antigorputzak, baina birusak berak "maltzurki agerian jarritako" molekula horiek baino ez ditu ezagutuko.
Bestela esanda, birus zaharrak edota zepa jakin batekoak erabiliz lortutako txertoek eskaintzen duten babesak ez du birus berrien edo beste zepatakoen kontra baliorik izangd. Eta noski, birusaren izaera edo "identitatea" ia urtero alda daiteke. jakina da bestalde, birusek ez dutela ezer apropos egiten. Beraien erreplikazio-tasa altua dela kausa, hautespen-prozesuek intentsitate handiz eragin ohi dute birusengan eta beren eboluzioa azlkarra izan ohi da. Eboluzio-prozesu horretan birus-motarik aldakorrenak izaten dira faboratutakoak, hauek sistema inmunitarioak gainditzeko erraztasun handiagoa izaten baitute.
Gauzak horrela, urtero Atlantan dagoen Munduko Gripe-Zentruko ikerlariak hurrengo gripe-izurria eragingo duen errudunaren identitatea asmatzen sutsuki saiatzen dira. Negua heldu baino bizpahiru hilabete lehenago, birusa identifikatu, purifikatu eta txertoa, kopuru handitan, prest izan behar du. Bestela, agure, gaixo kroniko eta bestelako inmunodeprimituek osatzen duten populazio-zati handi baten bizitza arriskuan legoke (zeharka bada ere, gripeak HIESak baino hildako gehiago eragiten omen du urtero). Gripearen kasuan, txertoa ez den beste irtenbide terapeutiko bat aurkitzeak duen garrantzia beraz, nabaria da.
Baina txertoen arrakasta-ezaren adibiderik ezagunena gaur zorigaiztoko ospea duen HIESaren agente birikoaren aurkako txertoarena dugu. GIB (Giza Inmunoeskasiaren Birusa) izeneko birus hori erretrobirus -motakoa da eta bere bizi-zikIoan, tarte luzea ematen du ostalari-zeluletan integraturik. Beraz, ostalarian sartu eta gero birusaren informazio genetikoa zelularen informazio genetikoaren artean ezkutatu eta bertan latentzi egoeran manten daiteke hainbat denboraz (egoera horrek hainbat urtez iraun dezakeela uste da).
Horrelako birusen kontrako defentsarik nagusiena infektaturiko zelulak detektatu eta suntsitzea da, hau da, inmunitate zelular deritzon prozesua burutzea. Askotan, zelula inbadituek agerian izaten dituzte beren azalean agente arrotzaren zatiak eta honela, nolabait esatearren, bere egoeraren berri ematen dute. Horrelakoetan, garbitze-funtzioa duten zelula espezializatuek "ikur" horiek detektatu eta zelula infektatuak suntsitu egiten dituzte. Zelula garbitzaileak, zelula T zitotoxikoak dira. Infekzio-egoeratan zelula hauek beroien hazkuntza bideratzen duten beste zelula bitartekarien beharra daukate.
Bitartekari espezializatuen funtzioa betetzen duten zelula horiek, linfozito laguritzaileak izaten dira; zehazkiago esanda, T4 motako linfozitoak. Gainera, linfozito horiek ezinbestekoak dira bestelako eraritzun inmunitarioak koordinatzeko. Tamalez, T4 linfozitoak GIB birusaren gogoko harrapakinak dira eta berez perfektua ez den defentsa-sistema hori akastunago bihur daiteke, GIBak eragindako infekzioaren ondorioz, bitartekari-lana egiten duten T4 linfozitoak desagertuz gero. Era honetara sistema kolapsora helduz gero, inmunoeskasia sortzen da eta ondorioz, organismoa babesikgabe geratzen da edozein infekzioren aurrean eta heriotza hel daiteke.
Infekzio-mota horrek, txertoaren garapena oztopatzen du. Batetik, txertoa eragin gutxiko sendabidea izango litzateke kutsatutako pertsonentzat, antigorputzek zelulatik at lan egiten dutenez kanpoko birus mihiztatuak (hau da, estalkia ere badutenak) baino ezingo baititu detektatu; latentzi egoeran dagoen GIB birusa detektaezina baita gure defentsentzako. Bestetik, orain artean eraginkorren izan diren txertoak birus ahulagoak erabiliz prestatutakoak izan dira, hau da, eskala txikiko infekzioak eragin ditzaketen txertoak. Bistakoa denez, pertsona osasuntsu bati ezin zaio horrelako GIB-txertorik eman, itzulezina eta hilkorra den infekzioa eragiteko arriskua handiegia baita.
Oztopo hori gainditzeko, txerto sintetikoak eta errekonbinanteak aztertzen ari dira gaur egun. Bi txerto-ereduak saiakuntza-fasean badaude ere, etorkizunean eraginkorrak izateko oztopo handi bat gainditu beharko dute: gripearen kasuan bezalaxe, GIBaren ezaugarri inmunogenikorik nabarmenenak aldakorrenak dira aldi berean, birusak berak nahita erakusten dituela diruditen horietakoak. Zati inmunogeniko aldaezinak zeintzuk diren bereizterik izango bagenu, txerto eraginkorra prestatzeko aukera gehiago izango genuke. Horretarako eta aurrerago ikusiko dugunez, birusaren biologia sakonki ikertu behar da. Bitartean eta hurbileko etorkizunean, GIBaren kontrako txertoak ametsa dirudi. 1994ko abunuaren hasieran Osakan egin zen Munduko HIES-Kongresura hurbildu ziren adituek horrela baieztatu zuten behintzat.
Gripe- eta HIES-pandemien arazoak gaixotasun horiek sortarazten dituzten birusen biologia ikertzera eramaten gaitu. Biak estalkidun birusak ditugu, hots, mintz zelularren moduko mintz lipidiko batez inguratuta daude (1. irudia). Izatez, birioiek mintza beraiek jaiotako zelularengandik lapurtzen dute. Dena dela, birusen mintzek ez dituzte zelularen mintzeko proteina berberak edukitzen, arantze proteina deritzen glikoproteina biriko bereziak baizik. Proteina espezifiko horietan egon liteke hain zuzen gripea eta GIBa bezalako birusen kontrako borrokaren koxka.
Gripe-birusaren sarrerari buruz munduko espezialistarik nagusiena den judith Whiteren hitzetan: " arantze proteinak ditugu sarrerarako funtsezko makinak, proteina balioanitz horiek sistema inmunitarioa engainatu, zelula-tropismoa (zein zelula mota inbadituko den) mugatzen duen hartzaile zelulorra ezagutu eta mintz zelular eta birikoa fusionatzearen funtzioak betetzeko goi baitira ". Aurrerago ikusiko dugunez, proteina horiek funtsezkoak dira prozesu horretan, aipatutako funtzio horietariko edozein blokeatuz gero, birioiek zeluletan sartzeko duten gaitasuna arriskuan egongo litzateke eta.
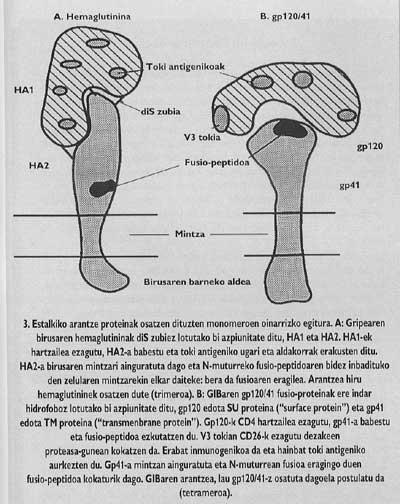
Sarrera gerarazten saiatzea metodo egokia izan liteke infekzio birikoaren aurkako borrokan. Horretarako ordea, arantze proteinen funtzioa eta egitura ondo ezagutu behar ditugu. Proteina horien zati funtzionalak zeintzuk diren ezagututa eta ostalarian sartzeko beharrezkoak dituzten faktoreak argiturik, sarrera gerarazteko bideak diseinatzen has gintezke. GIB eta gripearen birusen estalkiko arantze proteinak paradigmatikoak dira zentzu horretan. Horien egitura eta funtzioak dira gehien ikertu direnak (3-5. irudiak).
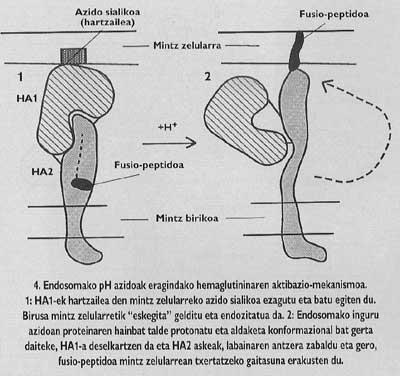
Mintz-proteina horiek egitura oligomerikoa dute, hau da, hainbat modulu desberdinez osatuta daude. Gripe-birusaren arantze proteina, hemaglutinina, heterotrimeroa da, hau da, bi azpiunitate desberdinez, HA1 eta HA2-z, osaturiko hiru monomeroz eraikita dago. Kapelako azpiunitatea HA1-a da eta fusio-peptidoa duen azpiunitatea HA2-a. Azken hori da endosomako pH azidoaz aktibatutakoan, fusio-peptidoa agertarazi eta helburu-zelularen mintzan txertatzen dena.
GIBaren arantze proteina tetrameroa da, hau da, gp120/41 izeneko lau monomeroz osatuta dago. Birus horretan, fusio-peptidoa duen azpiunitatearen (gp/41 izenekoa) aktibazioa gripearen birusaren zeharo desberdina da. Aktibazioa gerta dadin, azpiunitate hori kapelan dagoen gp120 izeneko azpiunitatetik askatu behar da iaz arte guztiz argi ez zegoen mekanismoa segiz, Gp41-a bi urratsetan askatzen da. Lehenean gp120-a linfozitoaren CD4 hartzailea ezagutu eta harekin bat egiten da. CD4-a ezagutzea ezinbestekoa da mintzen arteko fusio-prozesua hasteko, baina ez du azpiunitate fusogenikoa aktibatzen. lkerlariek ikusi zutenez, GlBak ez ditu infektatzen injinerutza genetikoz sortutako gizakiaren antzeko CD4 hartzailea duten arratoi-zelulak, nahiz eta zelula mutante horiek T4 linfozitoak adinako birusarekin bat egiteko gaitasuna erakutsi.
Hots, arratoi-zelula mutanteetako CD4 hartzaile horrek gizakiagan bezalaxe funtzionatzen zuen (birusari batzeko gai zen) arren, hartzailearekin bat egitea ez zen nahikoa fusioa bideratzeko. Beraz, CD4 hartzaileaz gain, beste faktore misteriotsuren batek eragin behar zuen gizakion T4 linfozitoen mintzetan, zelula horiek GIBarekiko sentikor izateko. Hori dela eta, GIBak bigarren hartzaile espezifikoren bat izan behar zuela postulatu zen orduan. 1993ko udazkenean, Pasteur Institutuko Ara Hovanessiaren taldeak hartzaile hori identifikatua zuela plazaratu zuen.
Aspalditik zen ezaguna Gp120-ren azalean dagoen V3 (3. irudia) toki antigenikoaren kontra sortutako antigorputz monokionalek infekzioa geraraz zezaketela. Gaur egun saiatzen ari diren gp120-an oinarritutako txerto sintetikoen balizko arrakasta toki hori ezagutzeko gai liratekeen antigorputzek eskain lezaketen babesaren menpe legoke. Toki horretan aminoazido-sekuentzia iraunkor bat dago, hots, belaunaldiz belaunaldi aldatzen ez den aminoazido-sekuentzia bat. Zergatik da berezia toki hori? Sekuentzia iraunkor hori "consensus" sekuentzia da. Proteasak proteinak zatitzen dituzten entzimak dira. Hala ere, entzimek ez dute nonahi mozten katea proteikoa, aminoazido-sekuentzia jakin batzuk dauden lekuetatik baizik. Sekuentzia horiei "consensus" sekuentzia esaten zaie.
Proteasa-mota bakoitzak "consensus" sekuentzia bakarra ezagutzen du eta ez besterik. V3-ko sekuentzia gizakiaren proteasa berezi batzuk baino ezin dute ezagutu eta horietariko bat T4 linfozitoaren azalean dagoen CD26 izeneko molekula hartzailea da. Pasteur Institutuko taldeak frogatu zuenez, ostalarian sartzeko, GIBak beharrezkoa du CD26-ren aktibitate proteasikoa. Horren ezean, ez dago infekziorik. Are gehiago, CD26-ren kontra ekoiztutako antigorputz monokionalak (CD26-rekin baino ezin direnak bat egin), edozein zelula-motan GIBaren sarrera saihesteko gai ziren. Gainera, normalean CD4-rik eta CD26-rik ez dauzkaten zelulek molekula horiek ekoiztu ditzaten eraginez gero, edozein zelula-mota GIBaz infektatzeko sentikor bihurtzea eragiten zuela aldarrikatu zuten talde horretako zientzilariek.
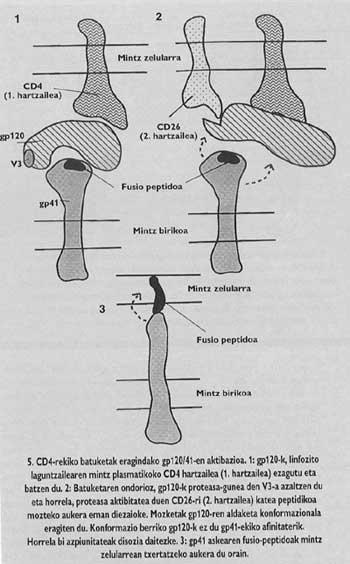
Horixe litzateke beraz, aipatu dugun faktore misteriotsua. Aurkikuntza horren arabera, gaur egun proposatzen den gp41-aren aktibazio-mekanismoaren bigarren pausuan, CD26-k moztu egingo luke katea proteikoa CD4-ri batuta legokeen gp120-ren V3 tokian. Mozketaren ondorioz, gp120-ren konformazioa aldatu egingo litzateke eta horrek gp41-ekiko afinitatea murriztea erakarriko luke. Afinitate-galerak, gp41-a helburu-zelularen mintzaren inguruan askatzea eragingo luke. Era horretan, fusio-peptidoa modu zuzenean txertatuko dela ziurtatzen da.
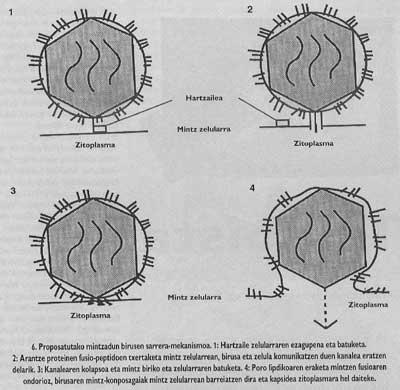
Sarreraren eskema orokorra osatu gabe dago oraindik. Hartzaileari batzea eta fusio-peptidoak txertatzea beharrezko pausuak direla jakin arren, oraindik ez dakigu birusaren eta helburu-zelularen mintzen arteko fusioa nola burutzen den. Hala ere, une honetan ditugun datu esperimentalek aditzera ematen digutenez, honako mekanismo ez-espezifiko hau izango litzateke (6. irudia) fusioa burutzearen kausa: fusio-peptidoak txertatu eta gero, fusioa eragin duen arantze proteinak birusa eta zelula komunika ditzakeen kanal bat eratzen du (pausu horrek oinarri esperimental handia duela esan dezakegu) eta ondoren, oraindik argi ez dagoen mekanismo bati esker kanaleak kolapsoa jasaten du eta birusaren mintza eta zelularena elkarren ondoan jartzen dira (pausu hori horrela gertatzen dela ziurtatzen duen esperimentu gutxi dago oraindik) eta, azkenik, mintzek bat: egitearen ondorioz, zelularen eta birusaren mintzetako lipido eta proteinak nahasi egiten dituen poro lipidiko bat eratzen da puntu jakin batetan (hau esperimentalki frogatuta dago). Poroa zabaldutakoan birusaren kapsidea zelularen zitopiasmara heltzeko aukera izaten du.
Oraindik ez dakigu prozesu orokor horiek zein neurritan izango zaizkigun baliogarriak birusen sarrera inhibitzeko, baina inhibizio hipotetiko hori gauzatzeko bide posibleak jorratuz joan gintezke. Halaber, prozesuaren oinarrizko mekanismoen xehetasunak argitu ahala, irtenbideak aurkitzeko aukera gero eta handiagoa izango da. Izan ere, deskribatutako sarrera-prozesua inhibitzen saiatzea ez da burutapen soila, gaur egun jadanik praktikan jartzen ari den errealitatea baizik. GIBaren infekzioa gerarazteko asmoz, CD4 hartzailearen zatiki soulgarriak erabiltzen ari dira jada terapia baten oinarri gisa.
Birioiak zatiki horiekin bat egin eta inaktibatu egingo direlakoan daude ikeriariak. Arestian aipatu dugunez, gp120-n oinarritutako hainbat txerto sintetiko saiatzen ari da peptido honen toki inmunologikoen kontrako antigorputzak GIBaren infekzioa gerarazteko gai izango direlako esperantzan. "In vitro" ikerketek sintesi kimikoz lortutako fusio-peptidoek GIBaren sarrera ekidin dezaketela erakutsi dute. Ez dakigu oraindik zein izan daitekeen inhibizio horren zergatia, baina baliteke peptido aske horiek distortsioak eragitea arantze proteinek eraikitako kanalen egituran. "In vitro" egindako esperimentuetan ikusi denez, V3-k dituen hainbat sekuentziaren aritzeko sekuentziak dituzten zenbait peptido sintetiko sarrera inhibitzeko gai dira.
ltxuraz, peptido horiek CD26-ren batze-prozesua gerarazteko gauza izan daitezke. Gripe-infekzioak tratatzeko asmoz, hemaglutininaren fusio-peptidoa askatzea inhibitzeko gai diren farmakoak diseinatzen ari dira. Farmako horiek HA2 azpiunitatearekin bat egingo lirateke, peptidoa kokatzen den gunetik hurbil, eta tapoi baten moduan, fusio-peptidoa mugitzea oztopatuko lukete. Egon badaude poro lipidikoak eratzea saihesteko gai diren agenteak, birusaren eta zelularen mintzak bat egitea oztopatzeko erabil litezkeenak, apika.
Birusaren sarrerari buruzko ikerketa hasi berria da. Etengabeko saiakera zientifikoan irteribide ugari agertuko dela pentsa daiteke. Naturak baimena ematen digun unean, pertsonengan erabili ahal izango den terapia antibirikoa diseinatzeko ordua helduko da. Terapia hori sarrera-prozesua inhibitzean izan lezake oinarria. Une hori iritsi bitartean, gaur egun argi dagoena zera da: infekzio birikoaren eremuan, txertoena bezalako ikerkuntzaren alderdi bakar batean iharduteak ez ditu irteribide posible guniak ekarriko. Sarrera birikoan parte hartzen duten oinarrizko mekanismo molekularrak ulertzeko ahaleginek ere eragin handia izango dute terapia posibleak bilatzeko orduan.

- birioi
Azido nukleiko (ADN zein ARN) eta estalki proteiko batez osatuta dagoen birus osoa. - erreplikazio
ADN molekula bat eredu gisa erabiliz beste bi molekula sintetizatzea. Zenbait kasutan, birusa beraren kopiak egitea. - genoma
Espezie bateko berezko geneen multzoa. Eukaritoetan ez bezala, prokariotoetan kromosoma bakar batean antolatua dago. - zepa
Ezaugarri genetiko eta patogeno berberak dituzten birusen multzoa. - antigeno
Organismora sartu ondoren antigorputzak sortzen dituen substantzia. - antigorputz
Antigenoa sartu ondoren organismoak sortzen duen substantzia (inmunoglobulina), inmunitate-mekanismoan parte hartzen duena. - erretrobirus
Material genetiko gisa ARNa duen birusa; mota honetako birus askok minbizia eragiten dute eta mota honetakoa da HIESa eragiten duena ere. - inmunitaten zelular
Linfozito (globulu zuri) eta makrofagoen bidez lortzen den inmunitatea. - glikoproteina
Bere osakeran aminoazidoez gain azukre-molekula bat ere baduen heteroproteina edo proteina konplexua. - inmunogeniko
Erantzun inmunea eragiten duena. - endozitatu
Zelula batek endozitosi deritzon prozesuaren bidez zelulaz kanpoko zerbait barneratzea. - endosoma
Zelula-nukleoaren osagai ez-desberdindua. - aminoazido
Karboxilo eta amino taldez osatutako molekula organikoa. Zenbait aminoazido proteinen osagai dira. - fusio-peptido
Birusaren mintza eta zelula-mintza bat egiteko prozesua martxan jartzen duen peptidoa. - peptido
Aminoazido-kopuru mugatu bat elkarri lotzen eratzen den molekula. - monomero
Bere berdinekin erreakziona dezakeen molekula. - monoklonal
Zelula-klon beretik sortutakoa. - konformazio
Atomo bati lotutako beste atomo edo erradikalen kokaera tridimentsionala.
Nieva, Jose Luis