



Estratègies del virus per a la invasió cel·lular
Els virus solen situar-se en el límit entre vius i inerts. Els virions no posseeixen maquinària metabòlica pròpia, és a dir, no són capaces de construir per si mateixos els seus components estructurals a partir d'antecedents metabòlics. Per això, per a poder reproduir-se és necessari envair a altres éssers (hostes) i apropiar-se de la maquinària metabòlica de les cèl·lules d'aquests.
Dins de la informació genètica que porten els virus protegits en el seu interior, s'acumulen les ordres per a inhibir el metabolisme de la cèl·lula infectada i orientar la síntesi dels components del virus. Per tant, els virus són paràsits moleculars que a vegades poden convertir-se en depredadors i portar a la mort sense pietat en ser infectat.
Per tant, per a replicar, els virus són imprescindibles que el seu genoma (ADN o ARN) entre a l'interior de l'hoste. El procés que han de seguir per a això es denomina introducció. Atès que l'entrada és obligatòria per a poder produir-se la lnección, això té gran importància en el cicle de vida dels virus. En el cas de virus que penetren en cèl·lules eucariotes, els virions han de superar la coberta molecular externa o membrana que envolta a la cèl·lula.
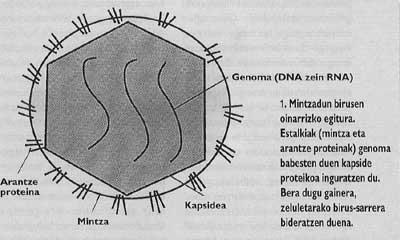
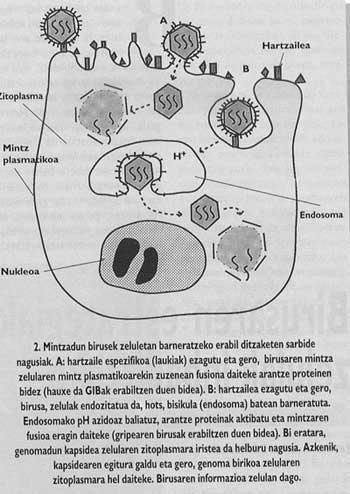
Així mateix, els animals i els virus més importants amb capacitat d'infectar a l'home estan envoltats d'una membrana com la de les cèl·lules anomenades cobertes (Figura 1). Amb la finalitat d'orientar l'entrada, aquests virus han desenvolupat mecanismes d'integració amb les cobertes de les cèl·lules que envairan les seves pròpies cobertes. Una vegada aconseguida la fusió de les membranes de la cèl·lula de destinació i el virió, el genoma del virus pot arribar al citoplasma de la cèl·lula (Figura 2) des d'on començar a replicar.
Les estratègies d'entrada desenvolupades pels virus encoberts són processos molt complexos que, per a comprendre'ls bé, els científics estan realitzant constants esforços, ja que si poguéssim aprendre com evitar l'entrada de l'enemic assassinat estaríem en vies de protecció per a certes infeccions. Amb l'objectiu de trobar una forma senzilla i global d'inhibir les infeccions víriques, els investigadors han buscat els elements comuns de l'entrada.
Desgraciadament, les coses no són tan senzilles com vulguem. Com sabem fins ara, almenys cada família de virus ha anat desenvolupant una estratègia d'entrada diferent, per la qual cosa la hipòtesi que existeix un mecanisme d'entrada general i comuna ha anat perdent força. A més, les recerques sobre els mecanismes d'entrada de virus encara no han donat molts resultats, ja que a principis dels anys 80, quan els científics es van adonar que no era possible produir una vacuna universal contra el virus que produeix la grip, aquest camp va començar a desenvolupar-se i pot considerar-se de recent creació.
En paraules de l'investigador espanyol Rafael Blasco: "la recerca sobre qualsevol virus està morta en el moment en què es desenvolupa la vacuna per a tractar la malaltia que provoca". El Sr. Blasco és l'especialista més important de l'Estat en la introducció de virus com el Poxvirus que crea la verola. El virus de Navarra és el primer ser que els éssers humans hem fet desaparèixer intencionadament del planeta, per la qual cosa Blasco diu que no hi ha massa responsabilitat sobre l'entrada dels seus familiars que es consideren poc importants.
L'error d'aquest plantejament s'ha posat de manifest en els últims temps, sense saber com detenir les plagues de grip i SIDA. Avui dia no és possible predir el tipus de virus que provocarà la següent epidèmia. Per a superar aquesta incapacitat, seria un pas important desenvolupar una teràpia universal que servís per a tractar qualsevol infecció vírica. Per a això també és necessari investigar en profunditat els processos d'entrada de virus. El virus de la grip, per exemple, és especialista en trufar amb la nostra protecció immunitària. Per a poder neutralitzar també els virus i altres agents estranys, les defenses del cos humà han de conèixer i diferenciar prèviament a aquests agents estranys. Això s'aconsegueix en l'actualitat mitjançant vacunes. Les vacunes posen de manifest la identitat dels agents estranys a les defenses del cos, de manera que s'aconsegueix que les cèl·lules que produiran anticossos que s'han d'unir a aquests agents estiguin llestes abans que es produeixi la infecció.
En el cas de la grip, no obstant això, les coses no són tan fàcils. Les bovines antigripales han de ser modificades anualment, ja que cada escòria del virus de la grip presenta antígens diferents (és a dir, diferents grups de molècules víriques que poden identificar els anticossos). És més, sembla que el propi virus canviarà en les següents generacions sabent que posa de manifest intencionadament aquestes molècules. D'aquesta forma podem produir anticossos enfront del virus amb facilitat, però només el propi virus coneixerà aquestes molècules "maliciosament exposades".
En altres paraules, la protecció que ofereixen les vacunes obtingudes mitjançant l'ús de virus antics o d'una escòria determinada no té valor enfront de nous virus o altres escòries. I per descomptat, la naturalesa o "identitat" del virus es pot canviar gairebé anualment. d'altra banda, és evident que els virus no fan gens propi. A causa de la seva elevada taxa de replicació, els processos selectius solen incidir amb gran intensitat en els virus i la seva evolució sol ser esclava. En aquest procés evolutiu els tipus de virus afavorits són els més variables, ja que aquests solen tenir una major facilitat per a superar els sistemes immunitaris.
Així les coses, els investigadors del Centre Mundial de Grip, situat cada any a Atlanta, tracten d'esbrinar amb entusiasme la identitat del responsable de la pròxima epidèmia de grip. Dos o tres mesos abans de l'arribada de l'hivern, el virus ha de ser identificat, purificat i preparat en grans quantitats. En cas contrari, una part important de la població formada per ancians, malalts crònics i altres immunodeprimits estaria en perill (indirectament, la grip provoca més morts anuals que la SIDA). En el cas de la grip, la importància de trobar una alternativa terapèutica diferent a la vacuna és evident.
Però l'exemple més conegut de la falta d'èxit de les vacunes és la vacuna contra l'agent víric de la SIDA, que avui gaudeix de fama desgraciada. Aquest virus, denominat VIH (Virus de la Immunodeficiència Humana), és un virus de tipus retrovirus que en el seu cicle de vida dóna un llarg espai integrat en les cèl·lules dels hostes. Per tant, després d'entrar en l'hoste, la informació genètica del virus pot ocultar-se entre la informació genètica de la cèl·lula i mantenir-se en estat latent durant diversos anys (s'estima que aquesta situació pot durar diversos anys).
La principal defensa contra aquesta mena de virus és la detecció i destrucció de cèl·lules infectades, és a dir, el procés denominat immunitat cel·lular. En moltes ocasions, les cèl·lules envaïdes presenten en la seva pell fragments d'agent estrany que, per dir-ho d'alguna manera, indiquen el seu estat. En aquests casos, les cèl·lules especialitzades amb funció de rentada detecten aquests "símbols" i destrueixen les cèl·lules infectades. Les cèl·lules netejadores són cèl·lules T citotòxiques. En situacions infeccioses aquestes cèl·lules necessiten d'altres cèl·lules intermediàries que permetin el creixement d'aquests.
Aquestes cèl·lules, que compleixen la funció d'intermediaris especialitzats, són limfòcits cedent, més concretament limfòcits tipus T4. A més, aquests limfòcits són imprescindibles per a coordinar altres homicidis immunitaris. Desgraciadament, els limfòcits T4 són preses favorites del virus del VIH i aquest sistema defensiu no perfecte en si mateix pot convertir-se en més defectuós si la infecció pel VIH provoca la desaparició dels limfòcits T4 que actuen com a intermediaris. Així, en arribar al col·lapse del sistema es produeix una immunodeficiència que fa que l'organisme quedi desprotegit davant qualsevol infecció i pugui arribar a la mort.
Aquest tipus d'infecció dificulta el desenvolupament de la vacuna. D'una banda, la vacuna seria una cura poc eficaç per a les persones infectades, ja que els anticossos només podran detectar virus assemblats externs al treball cel·lular (és a dir, amb coberta), ja que el virus VIH en latència és indetectable per a les nostres defenses. D'altra banda, les vacunes més efectives fins al moment han estat les preparades amb virus més febles, és a dir, vacunes que poden causar infeccions a petita escala. Òbviament, a una persona sana no se li pot administrar aquest tipus de vacuna de VIH, ja que el risc d'infecció irreversible i mortal és excessiu.
Per a superar aquesta barrera, actualment s'estan estudiant vacunes sintètiques i recombinants. Encara que tots dos models de vacunes estan en fase d'assaig, per a ser eficaços en el futur hauran de superar un gran obstacle: igual que en el cas de la grip, les característiques inmunogénicas més destacades del VIH són alhora les més canviants, de les que semblen mostrar intencionadament el propi virus. Si poguéssim diferenciar les parts inmunogénicas inalterables, tindríem més possibilitats de preparar una vacuna eficaç. Per a això i com veurem més endavant, és necessari investigar en profunditat la biologia del virus. Mentrestant i en un futur pròxim, la vacuna contra el VIH sembla un somni. Així ho van confirmar els experts que es van acostar al Congrés Mundial de SIDA que es va celebrar a Osaka a principis d'agost de 1994.
El problema de les pandèmies de grip i sida ens porta a investigar la biologia dels virus que provoquen aquestes malalties. Tots dos són virus amb coberta, és a dir, envoltats d'una membrana lipídica com les membranes cel·lulars (Figura 1). De fet, els virions roben la membrana de la cèl·lula que ells han nascut. No obstant això, les membranes dels virus no contenen les mateixes proteïnes que la membrana cel·lular, sinó les glicoproteïnes víriques especials anomenades proteïnes espinoses. En aquestes proteïnes específiques podria estar el coll de la lluita contra els virus com la grip i el VIH.
En paraules de judith White, el major especialista del món en la introducció del virus de la grip: "Les proteïnes d'espines són màquines essencials per a l'entrada, ja que aquestes proteïnes polivalents són les que enganyen al sistema immunitari, coneixen al receptor cel·lular que limita el tropisme cel·lular (quin tipus de cèl·lula s'envairà) i són superiors per a exercir les funcions de fusionar la membrana cel·lular i vírica". Com es veurà més endavant, aquestes proteïnes són essencials en aquest procés, ja que el bloqueig de qualsevol d'aquestes funcions podria posar en perill la capacitat de penetració cel·lular dels virions.
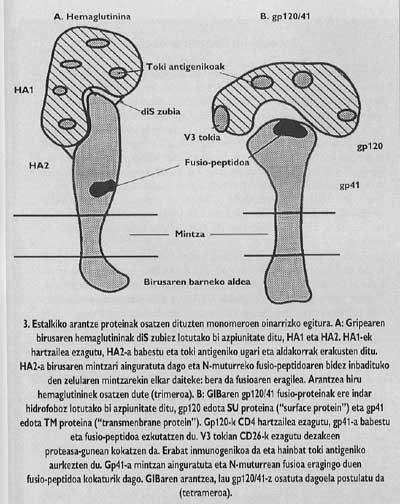
Intentar frenar l'entrada pot ser un bon mètode en la lluita contra la infecció vírica. Per a això necessitem un bon coneixement de la funció i estructura de les proteïnes espinoses. Una vegada conegudes les parts funcionals d'aquestes proteïnes i aclarits els factors necessaris per a la seva inclusió en l'hoste, podem començar a dissenyar vies que permetin la interrupció de l'entrada. Les proteïnes espinoses de la coberta del VIH i els virus de la grip són paradigmàtiques en aquest sentit. La seva estructura i funcions són les més investigades (Figures 3-5).
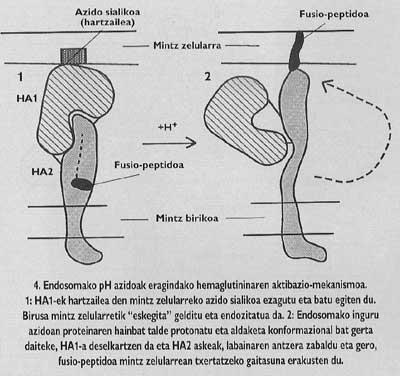
Aquestes proteïnes de membrana tenen una estructura oligómica, és a dir, estan formades per diferents mòduls. La proteïna espina del virus de la grip, l'hemaglutinina, és heterotrimero, és a dir, està construïda amb tres monòmers formats per dues subunitats diferents, HA1 i HA2. La subunitat de la capella és ha1 i la subunitat amb pèptid de fusió és ha2. Aquesta última és la que en activar-se amb el pH àcid de l'endosoma, fa aparèixer el pèptid de fusió i s'integra en la membrana de la cèl·lula de destinació.
La proteïna d'arç del VIH és tetràmer, és a dir, està formada per quatre monòmers anomenats gp120/41. En aquest virus, l'activació de la subunitat amb pèptid de fusió (anomenat gp/41) és totalment diferent al virus de la grip. Perquè es produeixi l'activació cal alliberar aquesta subunitat de la subunitat denominada gp120 que es troba en el barret, seguint un mecanisme que fins l'any passat no estava del tot clar, es deixa anar el Gp41 en dos passos. En la primera es coneix i s'integra el receptor CD4 del limfòcit gp120. El coneixement del CD4 és imprescindible per a iniciar el procés de fusió entre membranes, però no activa la subunitat fusogénica. Els lkerlaris van comprovar que GlBa no infecta cèl·lules de rates amb un receptor CD4 similar a l'humà generat per una enginyeria genètica, a pesar que aquestes cèl·lules mutants van demostrar la seva capacitat d'adhesió al virus equivalent al limfòcit T4.
És a dir, encara que aquest receptor CD4 de cèl·lules rates mutants funcionava com en l'ésser humà (era capaç d'unir-se al virus), la seva adhesió al receptor no era suficient per a canalitzar la fusió. Per tant, a més del receptor CD4, un altre factor misteriós havia d'influir en les membranes dels limfòcits T4 humans perquè aquestes cèl·lules fossin sensibles al VIH. Per això, es va postular que el VIH havia de tenir un segon receptor específic. A la tardor de 1993, l'equip d'Ara Hovanessia de l'Institut Pasteur va descobrir que havia identificat a aquest destinatari.
Antigament es coneixia que els anticossos monofuncionales creats contra el lloc antigènic V3 (Figura 3) en la superfície del Gp120 podien detenir la infecció. El possible èxit de les vacunes sintètiques basades en el GP120 que actualment s'està tractant dependria de la protecció que podrien oferir anticossos capaços de conèixer aquest lloc. En aquest lloc hi ha una seqüència permanent d'aminoàcids, és a dir, una seqüència d'aminoàcids que no canvia de generació en generació. Per què aquest lloc és especial? Aquesta seqüència permanent és la seqüència "consensus". Les proteasas són enzims que divideixen les proteïnes. No obstant això, els enzims no tallen per onsevulla la cadena proteica, sinó pels llocs on hi ha determinades seqüències d'aminoàcids. Aquestes seqüències es denominen consensus.
Cada tipus de proteasa només coneix una seqüència de consensus "". La seqüència en V3 només pot conèixer unes proteasas especials de l'ésser humà i una d'elles és la molècula receptora denominada CD26 que es troba en la superfície del limfòcit T4. L'equip de l'Institut Pasteur va demostrar que per a entrar en l'hostal, el VIH necessita l'activitat proteásica del CD26. En defecte d'això no hi ha infecció. És més, els anticossos monofuncionales fabricats contra el CD26 (que només es poden fusionar amb el CD26) eren capaços d'evitar l'entrada del VIH en qualsevol mena de cèl·lula. A més, si les cèl·lules que normalment no contenen CD4 ni CD26 provoquen la producció d'aquestes molècules, els científics d'aquest grup van reivindicar que qualsevol tipus de cèl·lula era sensible a la infecció per VIH.
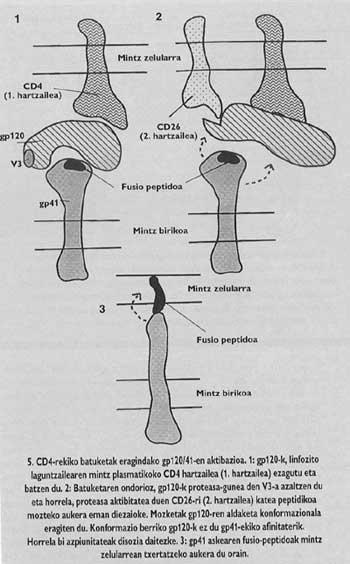
Aquest seria, per tant, el factor misteriós que hem esmentat. Segons aquesta troballa, en el segon pas del mecanisme d'activació del GP41 que es proposa en l'actualitat, el CD26 tallaria la cadena proteica en el lloc V3 del GP120 que estava unit a CD4. El tall suposaria un canvi en el conformat de GP120, la qual cosa suposaria una disminució de l'afinitat amb GP41. La pèrdua d'afinitat provocaria l'alliberament del gp41 al voltant de la membrana de la cèl·lula de destinació. D'aquesta forma s'assegura la correcta inserció del pèptid de fusió.
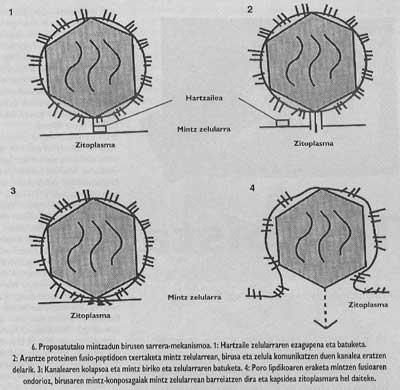
L'esquema general de l'entrada segueix sense completar-se. Encara que sabem que la unió al receptor i la introducció de pèptids de fusió són passos necessaris, encara no sabem com es realitza la fusió entre el virus i les membranes de la cèl·lula de destinació. No obstant això, les dades experimentals que tenim en aquest moment ens indiquen que el mecanisme inespecífic (Figura 6) seria el següent: una vegada inserits els pèptids de fusió, el virus de l'espina que ha produït la fusió forma un canal que pot comunicar la cèl·lula (aquest pas pot dir que té una gran base experimental) i posteriorment, gràcies a un mecanisme encara no clar, el canaló sofreix un col·lapse i, finalment, la membrana del virus i el lipídic. Una vegada obert el porus, la càpsida del virus pot arribar al citomopiámara de la cèl·lula.
Encara no sabem en quina mesura ens seran útils aquests processos generals per a inhibir l'entrada de virus, però podem anar treballant en possibles vies per a dur a terme aquesta hipotètica inhibició. Així mateix, a mesura que es vagin aclarint els detalls dels mecanismes bàsics del procés, la possibilitat de trobar solucions serà cada vegada major. De fet, intentar inhibir el procés d'entrada descrit no és una simple ocurrència, sinó una realitat que ja s'està posant en pràctica en l'actualitat. Amb l'objectiu de frenar la infecció del VIH, ja s'estan utilitzant les fraccions solubles del receptor CD4 com a base d'una teràpia.
Els investigadors creuen que els virions s'uniran a aquestes fraccions i s'inactivaran. Com ja s'ha esmentat anteriorment, diverses vacunes sintètiques basades en el GP120 intenten esperar que els anticossos contra els llocs immunològics d'aquest pèptid siguin capaços de frenar la infecció del VIH. Els estudis "in vitro" han demostrat que els pèptids de fusió obtinguts mitjançant síntesi química poden evitar l'entrada del VIH. No se sap encara quin pot ser la causa d'aquesta inhibició, però és possible que aquests pèptids lliures provoquin distorsions en l'estructura dels canals construïts per les proteïnes espinoses. En els experiments realitzats "in vitro" s'ha observat que diversos pèptids sintètics amb seqüències d'operació de diverses seqüències de V3 són capaces d'inhibir l'entrada de pèptids sintètics.
Amb l'ajuda de l'enciam, aquests pèptids poden ser capaços de frenar el procés de recollida del CD26. Amb l'objectiu de tractar infeccions gripals, s'estan dissenyant fàrmacs capaços d'inhibir l'alliberament de pèptid de fusió d'hemaglutina. Aquests fàrmacs s'unirien a la subunitat HA2, prop de la zona d'ubicació del pèptid, i com un tap, dificultarien el moviment del pèptid de fusió. Existeixen agents capaços d'evitar la formació de porus lipídics que poden ser utilitzats per a dificultar la fusió de les membranes del virus i de la cèl·lula.
La recerca sobre l'entrada del virus acaba de començar. Es pot pensar que en un assaig científic continu apareixeran molts punts d'inflexió. En el moment en què la naturalesa ens autoritzi, arriba l'hora de dissenyar una teràpia antiviral que es pugui utilitzar en les persones. Aquesta teràpia podria estar basada en la inhibició del procés d'entrada. Fins que arribi aquest moment, la qual cosa és clar avui dia és que en el camp de la infecció vírica, actuar en un sol aspecte de la recerca com la vacuna no va a possibles punts de tragèdia. Els esforços per comprendre els mecanismes moleculars bàsics que intervenen en la introducció vírica també tindran una gran influència en la cerca de possibles teràpies.

- tot un virus compost per un Àcid Nuclear (ADN i ARN) i una coberta proteica.
- síntesi d'altres dues molècules utilitzant com a model una molècula d'ADN de replicació. En alguns casos, fer còpies del propi virus.
- genoma Conjunt de gens propis d'una espècie. A diferència dels eucaríticos, en els procariotes està organitzat en un únic cromosoma.
- escòries Conjunt de virus amb les mateixes característiques genètiques i patògenes.
- substància que produeix anticossos després de la seva introducció en l'Organisme Antigen.
- substància (immunoglobulina) produïda per l'organisme després de la introducció de l'anticòs, que participa en el mecanisme d'immunitat.
- Retrovirus Virus que conté ARN com a material genètic, molts d'aquests tipus de virus provoquen càncer i d'aquest tipus és també el causant de la SIDA.
- immunitat obtinguda per immunitats cel·lulars linfocitas (glòbuls blancs) i macròfags.
- glicoproteïna En la seva composició, l'heteroproteína o proteïna complexa que conté, a més d'aminoàcids, una molècula de sucre.
- inmunogénico Responsable immune.
- endocitad Assimilació d'una cosa extracelul·lar a través del procés denominat endocitosi per una cèl·lula.
- endosoma Component no diferenciat del nucli cel·lular.
- molècula orgànica d'aminoàcids carboxil i grups aminos. Alguns aminoàcids són components de proteïnes.
- pèptid de fusió Pèptid que posa en marxa el procés d'elaboració d'una membrana de virus i una membrana cel·lular.
- molècula en la qual es produeix la unió d'un nombre limitat de pèptids Aminoàcids.
- monòmer Molècula capaç de reaccionar amb els seus iguals.
- monoclonal produït a partir del propi clon cel·lular.
- disposició tridimensional d'altres àtoms o radicals units a un àtom de conformat.
Buletina
Bidali zure helbide elektronikoa eta jaso asteroko buletina zure sarrera-ontzian







