



Orgànuls sense membrana
Quan a les escoles de biologia cel·lular s'expliquen les cèl·lules i les seves característiques, sempre s'esmenten dos tipus principals de cèl·lules: la cèl·lula eucariota i la cèl·lula procariota. I d'altra banda, quan s'enumeren les diferències entre tots dos tipus de cèl·lules, abans o després apareixen els orgànuls. Els orgànuls solen definir-se com a estructures especialitzades que compleixen funcions específiques a l'interior de la cèl·lula. En les explicacions sobre els orgànuls sempre s'ha esmentat el nucli, protector de la informació genètica cel·lular; els mitocondris i cloroplastos, productors d'energia; el reticle endoplasmàtic granulat i llis, que participen en la síntesi de lípids i proteïnes; i l'aparell Golgi, compost per vesícules de secreció que formen i distribueixen paquets de proteïnes. Totes elles tenen una funció específica i estan envoltades d'una membrana semipermeable.
En els últims 100 anys no hi ha hagut canvis en l'estructura cel·lular que coneixem. O sí? Al novembre de 2018, investigadors de l'Institut Sloan Kettering van publicar un nou òrgan en la revista Cell (Dt. & Mayr, 2018). Nou orgànul assignat a un domini TIGER (que explicarem més endavant)! Seríem davant un nou concepte que canviés la integritat de la cèl·lula?
En els últims anys, els investigadors han analitzat amb més deteniment la sopa d'aigua i molècules tant del citoplasma cel·lular com de l'interior dels orgànuls. Els investigadors han observat en el citoplasma i a l'interior dels orgànuls (és a dir, a l'interior dels orgànuls amb una densitat determinada envoltats de líquids d'una altra densitat) una gran quantitat de vesícules sòlides que, igual que els orgànuls membranosos, tenen funcions especialitzades. Aquestes vesícules sovint no es corresponen amb la definició d'un orgànul “clàssic”: d'una banda, aquests orgànuls no membranosos no tenen límits concrets, i per un altre, poden ser orgànuls efímers, poden reaccionar ràpidament sota la influència dels senyals químics i, al mateix temps, desorganitzar estructures especialitzades generades en la desaparició de l'estímul.
No obstant això, per la seva funció i importància han aconseguit l'atenció dels investigadors.
Hi ha diversos orgànuls sense membrana
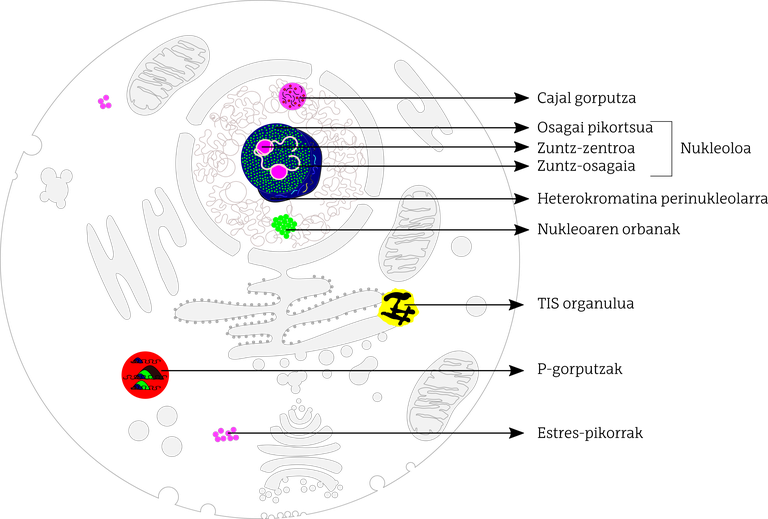
En les cèl·lules dels mamífers, els investigadors han identificat almenys una dotzena d'orgànuls sense membrana dispersos tant en el nucli cel·lular com en el citoplasma (Figura 1). En general, els orgànuls no membranosos que es troben dins del nucli estan especialitzats en el control de la transcripció de gens i en alguns aspectes del metabolisme de l'ARN. No obstant això, els orgànuls no membranosos dispersos pel citoplasma intervenen en el metabolisme, transport i homeòstasi de l'ARN missatger.
a) Orgànuls sense membrana dins del nucli
Entre els orgànuls no membranosos que es troben dins del nucli, destaca el nuclèol. El nuclèol és el més estudiat i probablement el més gran de tots. La seva funció principal és la biogénesis dels ribosomes necessaris per a sintetitzar proteïnes. En el nuclèol es distingeixen tres regions: centre de fibres, component fibrós dens i component granular. En la primera se situa l'ADN que codifica els ribosomes; la conversió a l'ARN d'aquest ADN, la transcripció, es produeix en el límit entre el centre de fibra i el component de fibra. En aquest límit apareix la proteïna coneguda com fibrilarina i es produeix el processament de l'ARN que codifica els ribosomes, l'adhesió al tall. El component granular, per part seva, és ric en proteïnes nucleofosmina, on es produeix l'acumulació i assemblatge de subunitats prerribosómicas. El nuclèol està envoltat d'ADN organitzat de forma molt compacta, a la qual es denomina heterocromatina perinucleolar. Són principalment rics en repeticions d'ADN, principalment satèl·lits d'ADN i conjunts d'ADN ribosòmics. Sembla que aquesta repetició de l'ADN del ribosoma evita la reconversió o recombinació de l'ADN, estabilitzant l'ADN ribosòmic.
Els cossos de Cajal també es troben en el nucli de la cèl·lula, però són més petits. Quant a l'estructura, estan constituïts per proteïnes i RNA que semblen ser necessaris per a l'assemblatge i modificació de snRNP (small nuclear ribonucleoprotein). Aquests snrnp són necessaris per als canvis posteriors a la transcripció del RNA missatger: entre altres, les parts transcrites d'RNA que no es convertiran en proteïnes, les intronas, les adhesions de tall. Els cossos de Cajal semblen participar en el procés de maduració final de les partícules snRNP i en la generació de complexos snRNP.
Les taques del nucli estan directament relacionades amb els cossos de Cajal i formen part del nucleoplasma. En les taques del nucli s'acumulen i modifiquen snRNP i proteïnes riques en aminoàcids serina i arginina. Tots ells participen en el procés de tall i pegat de l'ARN missatger.
b) Orgànuls sense membrana dispersos en citoplasma
Quant als orgànuls no membranosos dispersos pel citoplasma, generalment es denominen grànuls mRNP (messenger ribonucleoprotein). Encara que existeixen diversos tipus de RNP, és habitual que les proteïnes i l'ARN missatger es comparteixin i interactuin.
Els cossos de P (processing bodies) s'han descrit en diferents tipus de cèl·lules i s'ha observat que són àrees riques en proteïnes que intervenen en el transport, modificació i traducció de l'ARN missatger. Quan s'acumulen molts ARN missatgers no traduïts a aminoàcids, bé perquè la traducció està inhibida o en certes condicions d'estrès, augmenta la grandària i el nombre de cossos P.
Els grànuls d'estrès, com el seu nom suggereix, s'assemblen davant senyals d'estrès, segrestant molècules d'RNA missatger silenciades i factors de traducció. És a dir, la cèl·lula emmagatzema en zones concretes el material necessari per a fer front a aquest estrès. En els grànuls d'estrès és habitual trobar els factors necessaris per a iniciar la traducció i els components de les subunitats petites dels ribosomes.
Altres orgànuls citoplasmàtics sense membrana són els grànuls germinals, que apareixen en les cèl·lules germinals de l'embrió en desenvolupament (cèl·lules mare creadores de cèl·lules sexuals). Normalment són rics en ARN missatger i en enzims que modifiquen l'ARN. Sembla que intervenen en els canvis posteriors a la traducció de l'ARN missatger en les cèl·lules de la línia germinal.
Al setembre de 2018 van publicar en la revista Cell el descobriment de l'últim tipus d'orgànuls sense membrana: Va ser conegut com l'orgànul TIS. En interacció amb el reticle endoplasmàtic, permet interaccions entre proteïnes dirigides per l'extrem 3’UTR de l'ARN. La proteïna TIS11B està molt relacionada amb aquest orgànul, d'aquí el seu nom. La interacció entre l'orgànul TIS i el reticle es denomina domini TIGER. Aquest últim és diferent a la resta de mitjans del citoplasma, tant biofísics com bioquímicos. Tot això controlarà l'intercanvi de proteïnes del reticle endoplasmàtic, regulant les interaccions entre proteïnes, relacionant els extrems 3’UTR amb la proteïna TIS11B i organitzant una xarxa en el citoplasma i al voltant del reticle llis.
Relacionades amb malalties
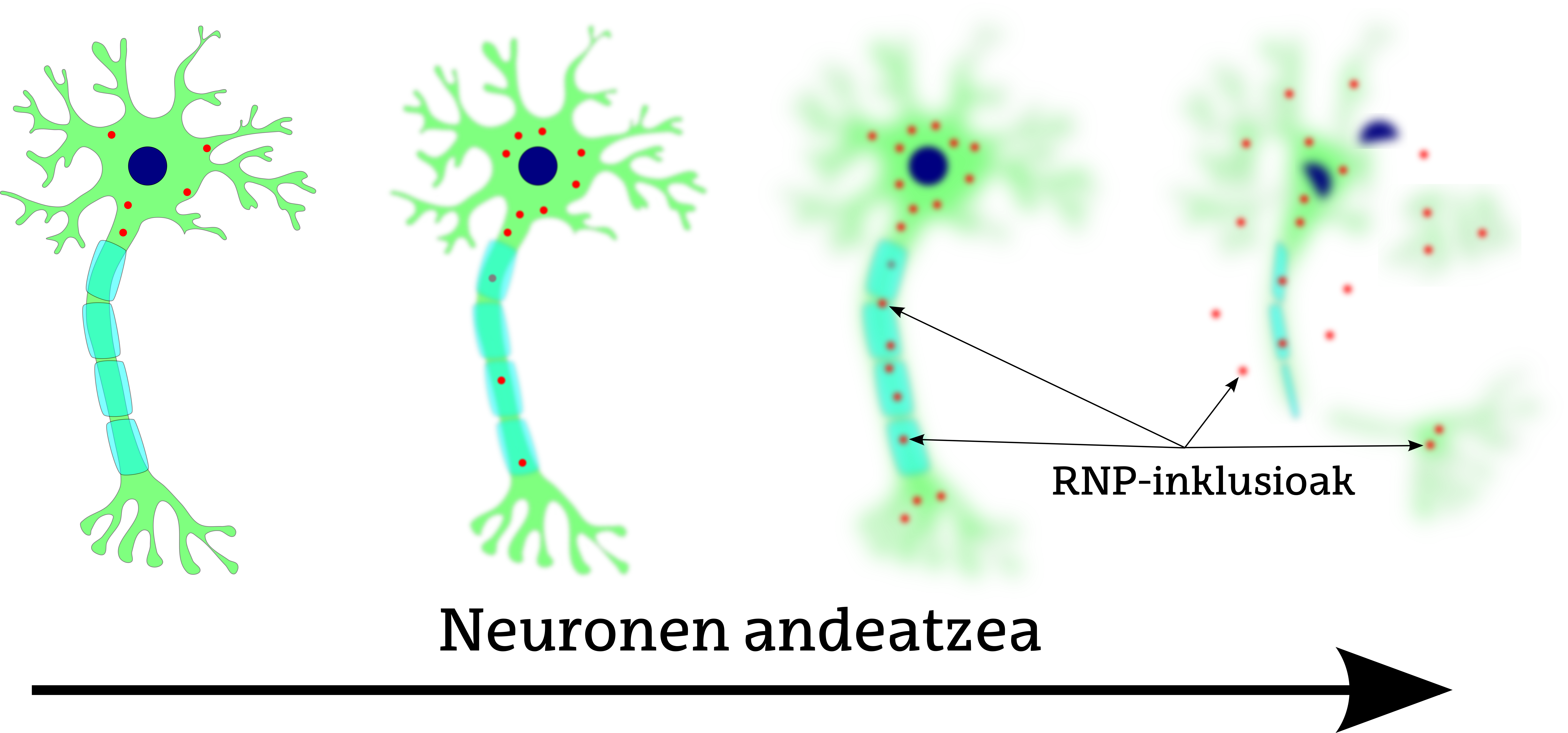
L'alt dinamisme dels orgànuls no membranosos fa que no sigui fàcil establir relacions directes entre els problemes de funcionalitat i les malalties d'aquests orgànuls. No obstant això, cal destacar el cas de l'esclerosi lateral amiotròfica. Una de les característiques d'aquesta malaltia (com altres malalties neurodegeneratives) és que es produeixen inclusions de missatgers de ribonucleoproteínas (RNP) que alteren el metabolisme normal de l'ARN missatger (Figura 2). Aquestes inclusions patològiques estan formades per grànuls proteics RNP, la gran acumulació dels quals produeix un assemblatge entre ells i la formació de fibres amiloides. Les fibres amiloides alteren el funcionament normal de les cèl·lules, degradant-les.
En els casos de càncer, els orgànuls no membranosos també presenten interessants opcions o perspectives. Un dels factors que regeixen el creixement cel·lular és l'assemblatge de ribosomes, que com s'ha vist té una gran importància en la producció de ribosomes. El nuclèol, per tant, pot ser una important destinació dels tractaments contra el càncer, caracteritzat pel creixement incontrolat del nuclèol i l'augment del nuclèol.
La clau del naixement de la vida? Repensant Oparin
En 1922, el bioquímic rus Alexander Oparin va publicar una teoria sobre l'origen de la vida en els inicis de la història de la Terra. Segons la seva teoria, es van formar els primers compostos orgànics simples, ja que l'energia elèctrica dels raigs o l'energia calorífica dels volcans va provocar la reacció del metà, l'amoni, l'aigua i altres components, rics en l'atmosfera reductora de l'època. Tots aquests ingredients es van organitzar en petites gotes denominades coacerbatos. L'any 2016 un grup d'investigadors alemanys van descriure els actius químicament i les gotes de líquid amb capacitat de fragmentació i, per tant, de perpetuació. Aquestes gotes i els orgànuls no membranosos tenen algunes característiques iguals, per la qual cosa existiran claus per a explicar l'origen de la vida en els orgànuls sense membrana?
Referències
Crabtree, M., Nott, T. 2018. These organelles have no membrane. The Scientist. Desembre 2018. https://www.the-scientist.com/infographics/infographic--what-are-membraneless-organelles--65135
Gómez, E., Shorter, J. 2018. "The molecular language of membraneless organelles". Journal of Biological Chemistry 1-23.
Dt., W., Mayr, C. A. 2018. "Membraneless Organelle Associated with the Endoplasmic Reticulum Enables 3< UTR-Mediated Protein-Protein Interactions". Cell 175: 1492-1506.
Mao, I.S., Zhang, B., Spector, D.L. 2011. 'Biogénesis and Function of Nuclear Bodies'. Trends in Genetics 27: 295-306.
Mitrea, D.M., Kriwacki, R.W. 2016 "Phase separation in biology; functional organization of a higher order". Cell Communication and Signalling 14: 1.
Buletina
Bidali zure helbide elektronikoa eta jaso asteroko buletina zure sarrera-ontzian







