Les bactéries ont également squelette
Chiots confus. Jusqu'à récemment les bactéries étaient dans les yeux des biologistes : sacs contenant du matériel génétique, quelques enzymes et autres molécules mélangées ; sans structures stables, sans ordre, sans squelettes. Mais les choses changent. Dans ces sacs ont été trouvés différentes structures. Beaucoup d'entre eux sont équivalents aux composants du cytosquelette des cellules eucariotes.
Dans les cellules eucariotes, c'est-à-dire des plantes, des animaux, des champignons et des protistes, on connaît bien le tissu intérieur appelé cytosquelette. Il s'agit d'une structure dynamique, composée de trois types de composants : microfilaments (actine), filaments moyens et microtubules (tubuline). Et il remplit plusieurs fonctions : donner et maintenir la forme aux cellules, faciliter le mouvement, le transport intracellulaire et la division cellulaire...
On considérait que les bactéries n'avaient pas de cytosquelette, ce qui était considéré comme le développement après la formation d'eucariotes (provenant de bactéries). Mais maintenant, il semble que ce n'était pas le cas: l'origine des actines, des filaments moyens et des tubulines pourrait être dans les bactéries.
Les bactéries sont passées d'être des protagonistes secondaires à être à la mode dans le domaine de la biologie cellulaire, et les découvertes se succèdent.
Derrière les formes

On peut dire qu'en 2001 la roue des découvertes a pris le rythme. Cette année-là, un groupe de chercheurs de l'Université de Newcastle en Angleterre a obtenu pour la première fois de voir la structure de la protéine MreB dans la bactérie Bacillus subtilis: Les fibres MreB forment une hélice sous le mur des bactéries cylindriques.
La protéine MreB est très semblable à l'actine des eucariotes et peut avoir la même origine. En outre, ils remplissent une fonction similaire. Dans les bactéries cylindriques les protéines MreB et très proches d'elle ont une fonction structurelle, c'est-à-dire, donnent forme à la bactérie.
En fait, les chercheurs ont vu que si elles manquent cette protéine change son apparence. B. subtilis, le MreB affecte la largeur de la bactérie et le Mbl à la longueur ; et E. Les bactéries coli deviennent cylindriques à sphériques lorsque le MreB leur est enlevé.
En fait, les bactéries sphériques n'ont pas MreB. De plus, en analysant les génomes de différentes bactéries et arcs (les deux groupes de procaryotes), les chercheurs de Newcastle ont découvert que les procaryotes cylindriques, filamenteux et hélicoïdaux contiennent un ou plusieurs gènes de protéine MreB, tandis que les sphériques en ont aucun.
En outre, dans des formes plus complexes de certaines bactéries, il est très probable que le cytosquelette est lié. Pour l'instant, vous n'avez trouvé qu'un exemple : Les bactéries Caulobacter crescentus ont une forme de haricots, et il semble que l'autre protéine leur donne cette courbure. En enlevant cette protéine, ces bactéries prennent une forme cylindrique. Cette protéine est krescentine et est très semblable au filament moyen des caryotes.
Clés de la division
En dehors des formes, le cytosquelette a également une grande importance dans la division cellulaire. Et là, les procaryotes n'ont rien en commun. Les bactéries présentes dans le milieu approprié peuvent être divisées toutes les demi-heures. Car une des clés de cette efficacité peut être précisément dans le cytosquelette. Mais les scientifiques n'ont fait que commencer à clarifier les mécanismes.
La protéine FtsZ est l'un des protagonistes de la division cellulaire des procaryotes. Forme un anneau dans la zone où la division est effectuée, au centre de la cellule. En enlevant cette protéine aux bacilles cylindriques, elles sont étirées et étirées sans se diviser en deux. De leur côté, les Staphylococcus aureus sphériques, une fois éliminé le gène de FtsZ, commencent à construire la nouvelle paroi cellulaire n'importe où et peuvent augmenter le volume huit fois avant qu'il explose.
En quelque sorte, la protéine FtsZ contrôle où il faut construire le nouveau mur. Autour de lui, il recueille les protéines qui construisent le mur. Dans le cas de S. aureus forme l'anneau à l'équateur et gouverne la création des deux nouveaux hémisphères. Dans les bacilles, pour sa part, il semble qu'au début la protéine MreB provoque l'étirement de la cellule, puis entre en jeu le FtsZ, créant de nouvelles extrémités arrondies pour diviser la cellule en deux.
Les anneaux de FtsZ peuvent avoir une grande importance à l'origine de la division cellulaire. Un groupe de chercheurs de l'Université Duke de Caroline du Nord a récemment vu que cette protéine forme également des anneaux dans les gouttes microscopiques d'huile. Maintenant, bien que la division cellulaire des bactéries est un mécanisme plus complexe, les chercheurs croient que les cellules les plus primitives auraient assez FtsZ pour se diviser en deux.
D'autre part, quelques organites des cellules eucariotes, les mitochondries et les chloroplastes emploient l'anneau de FtsZ pour se diviser. Cela renforce, en définitive, la croyance que ces organites sont des procaryotes qui, à leur époque, ont été introduits dans une cellule eucariote.
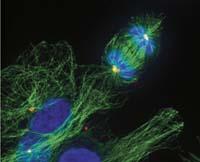
Outils modifiés
Les eucariotes ont également une sorte de protéine FtsZ, la tubuline. Et la tubuline participe aussi à la division cellulaire, mais elle remplit une fonction très différente : elle maintient les chromosomes correspondant à chaque nouvelle cellule pendant la division. Cela garantit que chaque nouvelle cellule aura ses chromosomes.
Il est curieux que ces deux protéines d'une même origine remplissent une fonction si différente. C'est-à-dire, FtsZ et la tubuline sont deux versions d'un même outil que les procaryotes et eucariotes utilisent pour effectuer des travaux très différents. Pour le même travail, ils utilisent des outils très différents. D'une part, pour le travail de la protéine FtsZ, équivalent à la tubuline, les eucariotes utilisent un outil d'actine. Et d'autre part, les procaryotes font ce que les caryotes font avec la tubuline avec une protéine équivalente à l'actine.
Et c'est que, comme les eucariotes, les bactéries doivent également bien distribuer le matériel génétique dans la fragmentation. Les bactéries possèdent un chromosome unique, ainsi que des anneaux d'ADN appelés plasmides. Les plasmides apportent une résistance aux antibiotiques et autres «extra» aux bactéries, il est donc très important que les nouvelles bactéries restent avec une copie des plasmides.
Comme les tubulines avec les chromosomes des eucariotes, la protéine ParM produit la séparation de certains plasmides, en éloignant les copies. Et ce ParM est une version bactérienne de l'actine.
ParM est une chaîne de protéines avec une capacité d'auto-addition et d'élimination des unités. La chaîne ParM rejoint les deux nouvelles copies du plasmide à distribuer. Par la suite, en ajoutant de nouvelles unités commence l'étirement de la chaîne, qui envoie les deux plasmides aux deux extrémités de la cellule.
C'est le mécanisme de distribution de certains plasmides. Mais qu'en est-il du chromosome bactérien? Comment distribuent-ils les copies du chromosome ? Cela reste un mystère.
Et ce n'est pas la seule. Il reste un long chemin à parcourir pour bien comprendre l'organisation et le fonctionnement des cellules des procaryotes. En plus de ceux déjà mentionnés, ils ont trouvé beaucoup plus d'éléments du cytosquelette et sont trouvés. Aussi des protéines qui n'ont rien dans le cytosquelette des eucariotes. Mais nous savons peu des mécanismes qui sont derrière tous ces éléments. Il reste encore beaucoup à faire dans ces chiots.
Buletina
Bidali zure helbide elektronikoa eta jaso asteroko buletina zure sarrera-ontzian